藍藻とは
藍藻(らんそう)とは文字通り藍色の藻類のことで、他の藻類や陸上の植物と同じように、太陽エネルギーによって光合成を行う独立栄養生物です。しかし主要な光合成色素は、クロロフィルaとbを持つ被子植物などと異なり、クロロフィルaとフィコビリン類(フィコエリトリンとフィコシアニン)です。種類や生育場所によって色素組成が異なるため、藻体の色は藍色から赤色、青色、緑色、緑褐色、または黒色に近いものまで様々です。分類の項目で詳しく述べますが、藍藻の体制は単純で、単細胞か群体を形成するか、あるいは糸状体です。また、有性生殖は知られていません。
さらに、藍藻は核という構造を持たない原核生物であるという点で、他の藻類や陸上植物と大きく異なります。そこで藍藻は原核生物の細菌類と同じ仲間と見なされ、最近ではシアノバクテリア、または藍色細菌と呼ばれることが多くなっています。一方、藍藻細胞と葉緑体は構造的にも機能的にもよく対応することから、藍藻は葉緑体の起源ではないかと考えられています。
藍藻は海や陸の水域だけではなく、地表面や樹上にも広く生育します。また寒帯から温帯、熱帯まで、極地や高山から温泉にまで、非常に様々な環境に広く生育しており、他の生物と共生しているものもあります。
33~35億年前の地層から、現在の藍藻に似た微化石が発見されていることから、藍藻の祖先は地球の酸素の生みの親と考えられています。地質時代の早期に出現した藍藻は、その形態をほとんど変えることなく生活域を広げ、今日まで生き続けています。その姿はまさに“生きた化石”と呼ぶにふさわしいものです。
藍藻の分類
「淡水浮遊性藍藻データベース」では、これまでに日本で報告された淡水浮遊性藍藻の様々な属の特徴と、よく観察される種、大発生が記録された種などを中心とした各種の形態の特徴と同定のポイントを記述します。
藍藻類の分類は主に藻体や細胞の形態的な特徴を比較し、それらの差異によって分類されてきました。しかし現在では、そのような形態的特徴だけでなく、藻体の生態的特徴や生理的特徴の比較、遺伝子情報の解析
(特に16S rRNA遺伝子、rbcL遺伝子、16S-23S rRNA ITS領域およびその二次構造)、透過電子顕微鏡を用いた細胞内のチラコイド配列の解析など、多面的手法を用いる必要があると認識されています。
2000年代に入ってからは特に、このような多面的手法によって多くの新種、新属が続々と発表されました。その結果、旧来の分類体系を再考する必要が出てきました。
そこで2014年に、それまでに発表された知見をもとに、藍藻類を8つの目に分け、それぞれの目に含まれる科と属がまとめられました(Komárek et al. 2014*)。
以下にこれら8つの目の特徴を示し、合わせて各目における日本で報告のある浮遊性藍藻の属を解説します。藍藻類の分類学的研究は現在も進行中で、本データベースに示すものが最終的な見解というわけではありません。研究の進展状況に応じて本データベースの改定を行う予定です。
*Komárek et al. 2014. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 86: 295-335.
グロエオバクテル目 Gloeobacteriales
チラコイドをもたない藍藻グループ。淡水浮遊性藍藻データベースに本目は含まれていない。
シネココックス目 Synechococcales**
単細胞であるか多数の細胞が規則的または不規則的に集まって群体を形成するグループ(グループA)と、細胞が一列に密着して並ぶ糸状体を形成するグループ(グループB)の2つが含まれている。 いずれの場合も栄養細胞のみで構成される。糸状体を形成するグループではトリコームの幅は狭く、多くは3µm以下で、真分枝はない。どちらのグループもチラコイドは細胞の周縁部にある。
A. 球形、楕円形、または円筒形の細胞が単独で生育するか、または細胞が集合して群体を形成する属
アナテケ属 Anathece
透明な粘質内に多数の楕円形ないし円筒形の細胞が集合して群体を形成する。群体は非常に微小で浮遊性。細胞の幅は約1μmほどでガス胞はない、粘質鞘をもたない。
本属はアファノテケ属(Aphanothece)の亜属から階級変更された。アファノテケ属は群体や細胞の形態が本属とよく似ているが、群体は肉眼でわかる程度に大きくなり、
底生、着生、気生または一時浮遊性で、個々の細胞が粘質鞘をもち、細胞の幅は1μm以上とやや大きい点でアナテケ属と区別できる。
また、チラコイドの配列および遺伝子解析の結果からも両属は区別できることが分かっている。
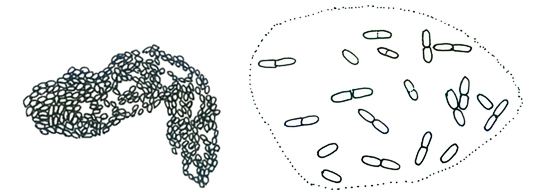
アファノカプサ属 Aphanocapsa
透明な粘質内に多数の球形の細胞が集合して群体を形成する。群体は浮遊性。個々の細胞は粘質鞘をもたない、ガス胞はない。 本属はミクロキスティス属(Microcystis)と同様に群体は立体的に広がるが、細胞がガス胞をもたない点で区別できる。
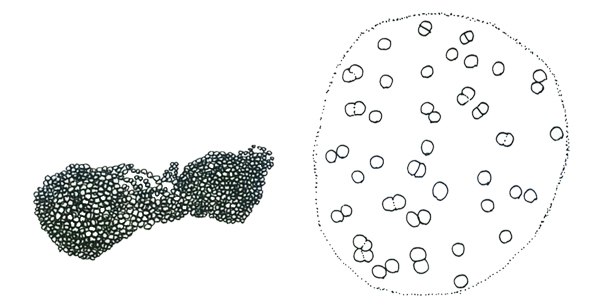
コエロスファエリウム属 Coelosphaerium
球形の透明な粘質鞘の表層に、ほぼ球形の細胞が一層に等間隔に並び、浮遊性の群体を形成する。群体の内部に糸状や紐状の構造物はない。 個々の細胞は粘質鞘をもたない。
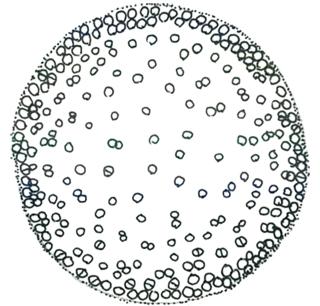
キアノディクティオン属 Cyanodictyon
紐状の粘質内に多数の細胞が一列に連なって群体を形成する。藻体は浮遊性。紐状の群体は折れ曲がるように平面または立体的に広がり、 球状や網目状になる。個々の細胞は粘質鞘をもたない、ガス胞はない。

キアノグラニス属 Cyanogranis
透明な粘質内に少数ないし多数の細胞が密接に集合して群体を形成する。群体は浮遊性。群体の周縁部にある数個の細胞に鉄化合物が沈着している。個々の細胞は粘質鞘をもたない、ガス胞はない。

レンメルマンニエラ属 Lemmermanniella
球形の透明な粘質鞘の表層にほぼ円筒形の細胞が接線方向に一層に並び、浮遊性の群体を形成する。群体の内部に糸状や紐状の構造物はない。個々の細胞は粘質鞘をもたない、ガス胞はない。
リムノコックス属 Limnococcus
透明な粘質内に少数の球形または分裂直後の半球形の細胞が集合して群体を形成する。群体は浮遊性。個々の細胞は粘質鞘をもたない、ガス胞はない。
本属は群体形態の差異、個々の細胞が粘質鞘をもたないこと、細胞分裂の違い、さらに遺伝情報の解析結果からクロオコックス属(Chroococcus)と区別できるとして、クロオコックス属の亜属から階級変更された。
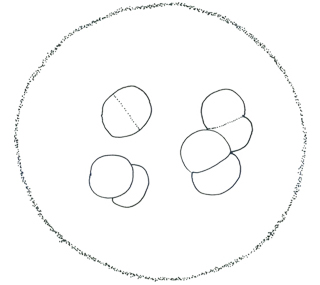
メリスモペディア属 Merismopedia
数個ないし多数の細胞が一層に規則正しく縦横に並び、平板状の群体を形成する。浮遊性。

パンヌス属 Pannus
透明な粘質内に球形の細胞が1-2(-3)層に平板状に並び、浮遊性群体を形成する。群体の形態は円形、葉状、網目状など。個々の細胞は粘質鞘をもたない。
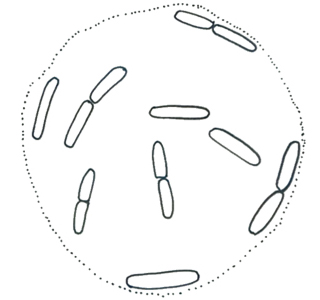
ロメリア属 Romeria
藻体は幅1-3µmの細胞が縦一列に数個ないし30個ほどが折れ曲がりながら連なっている。細胞は円筒形または様々な程度に湾曲する、ガス胞はない。細胞先端は丸みを帯びている。鞘はないが透明な粘質に覆われていることがある。
スノウェラ属 Snowella
ほぼ球形の透明な粘質鞘の表面に細胞が一層に並び、浮遊性の群体を形成する。細胞は群体中央部から二叉分岐しながら外側に向かって放射状に伸びる粘質糸の先についている。細胞が2個または4個ずつ密接して並んでいることが多い。個々の細胞は粘質鞘をもたない。
ウォロニチニア属 Woronichinia
球形、卵形または小群体が密着した不規則な形の群体を形成し、浮遊する。細胞は球形、長球形または広卵形。群体の中心から放射状に並ぶ粘質の柄を持ち、柄の先端部である群体表面近くに細胞が並ぶなどの点でMicrocystisと区別できる。

B. 細胞が連結して糸状に並び、トリコームを形成している属およびトリコームの周囲に粘質鞘が密着している属
プランクトリングビア属 Planktolyngbya
藻体は一本のトリコームと、明瞭な粘質鞘からなる細い糸状体で、単独で浮遊する、運動性がない。トリコームは幅4μm以下、全長がほぼ同じ幅で、まっすぐまたはらせん形、先端は丸く終わる、カリプトラはない。細胞はガス胞をもたない。

プセウドアナベナ属 Pseudanabaena**
藻体は幅1-3μmの細いトリコームからなる。トリコームは全長がほぼ同じ幅で、先端は丸く終わる、カリプトラはない。細胞は長短の円筒形、細胞間の隔壁部でくびれる。
**Komárek et al. (2020)は、 シネココックス目Synechococcalesを、プセウドアナベナ目Pseudanabaenales 、シネココックス目およびサーモスティクス目Thermostichalesの3目に分けるという見解を示している。 これによると、Pseudanabana属はプセウドアナベナ目に含まれる。プセウドアナベナ目にはこの他にGeitlerinema属、Limnothrix属などが含まれるが、「淡水浮遊性藍藻データベース」にはPseudanabana属以外は記載されていない。
**Komárek et al. 2020. Phylogeny and taxonomy of Synechococcus-like cyanobacteria. Fottea 20(2):171-191.
スピルリナ目 Sprulinales
細胞は一列に並びトリコームを形成する。トリコームは細い規則的なラセン形を示し、栄養細胞のみで構成され、鞘はない。チラコイドは細胞周縁部にある。
スピルリナ目は現在のところスピルリナ科SpirulinaceaeのGlaucospira属、Halospira属、Spirulina属の3属が知られている。これらはすべてトリコームが規則的なラセン形を示す。
浮遊性で淡水域に生育するものはGlaucospira属だけである。Halospira属は高塩性水域に生育する。Spirulina属は基質上に多数のトリコームが集合して付着し、薄いマット状の塊となることが多く、淡水域だけでなく汽水、塩水、温泉などにも生育する。
「スピルリナ」の商標名で知られているものはスピルリナ目の種ではなく、ユレモ目の種である。
グラウコスピラ属 Glaucospira
トリコームは細く(3μm未満)、短く、規則正しい緩いらせん状で、単独で浮遊する。トリコームは全長が同じ太さで、鞘をもたない。細胞隔壁部でくびれない。細胞はガス胞をもたない。

クロオコックス目 Chroococcales
球状の細胞が単独で生育するか、または多数の細胞が規則的または不規則的に集まって群体を形成する。栄養細胞のみで構成される。チラコイドは細胞内に不規則に配列する。
クロオコックス属 Chroococcus
粘質鞘内に1個または少数の細胞が集合して群体を形成する。藻体は浮遊性または着生。粘質鞘は層状で細胞または細胞群のすぐ外側を覆っている。細胞にガス胞はない。
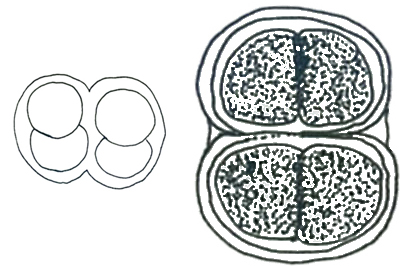
ミクロキスティス属 Microcystis
透明な粘質内に多数の球形の細胞が集合して浮遊性の群体を形成する。群体の粘質の外縁は不明瞭でほとんど見えないものと明瞭なものがある。個々の細胞はガス胞をもつ、粘質鞘をもたない。
ミクロキスティス属の藻は関東から九州、沖縄にいたる地域で発生するアオコの主な構成要素で、ミクロキスチンを産生するので、水源の湖沼ではその挙動が注意深く監視されている。2種以上混生することが多く、その組み合わせや相対的な量は時とともに変化する。
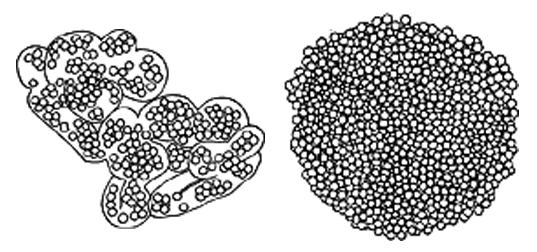
以下にミクロキスティス属(Microcystis)の日本でみられる種について、形態の比較一覧表を示す。
▼表中の学名をクリックすると詳細をご覧になれます。
| 種名 | 群体の大きさ | 群体の形態 | 群体外縁 | 細胞の大きさ | 細胞密度 |
|---|---|---|---|---|---|
| M. aeruginosa | 大型化する | 網状、帯状、楕円状など多様 | 不明瞭 | 3μm以上 | 密 |
| M. firma | 小さい | ほぼ球形 | 不明瞭 | 3μm以下 | 密 |
| M. flos-aquae | 小さい | ほぼ球形 | 不明瞭 | 3μm以上 | 非常に密 |
| M. ichthyoblabe | 大型化する | 大小のいびつな球形、スポンジ状 | 不明瞭 | 3μm以下 | やや疎 |
| M. novacekii | 小さい | 小型群体が緩く集合 | 不明瞭 | 3μm以上 | 密 |
| M. smithii | 小さい | ほぼ球形 | 不明瞭 | 3μm以上 | 疎 |
| M. viridis | 比較的小さい | 立方体の小群体が密に集合 | 不明瞭 | 3μm以上 | 8細胞の立方体を形成 |
| M. wesenbergii | 大型化する | 球状、帯状、楕円状など多様 | 明瞭 | 3μm以上 | やや疎 |
ラディオキスティス属 Radiocystis
透明な粘質内に細胞が放射状に配列した浮遊性群体を形成する。個々の細胞は粘質鞘をもたない。
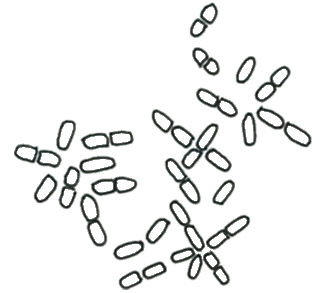
プレウロカプサ目 Pleurocapsales
単細胞、群体、偽柔組織を形成するなど、藻体の形態は多様だが、いずれも付着性という特徴がある。淡水浮遊性藍藻データベースに本目は含まれていない。
クロオコッキディオプシス目 Chroococcidiopsidales
水中や陸上の石などに付着し、岩内や砂漠などでも見つかっている。淡水浮遊性藍藻データベースに本目は含まれていない。
ユレモ目 Oscillatoriales
細胞は一列に並びトリコームを形成する。トリコームは幅がほぼ4μm以上多くは7μm以上で、栄養細胞のみで構成され、分岐しない。チラコイドは放射状、束状、または不規則に並ぶ。
アナグノスティディネマ属 Anagnostidinema
藻体は幅1-3μmの細いトリコームからなる、鞘をもたない、運動性があり、縦軸方向に滑走運動をする、先端部を曲げながらまたは回転しながら運動することもある。
トリコームの先端は細くなることはあるが常に丸みを帯びて終わり、乳頭状になることはない、尖ることはない、カリプトラはない。
本属と同じく滑走運動するガイトレリネマ属 Geitlerinemaは、トリコーム先端細胞が細長く伸びて曲がり、乳頭状に終わることで区別できる。
また、チラコイドの配列および遺伝子解析の結果からも両属は区別できることが分かっている。
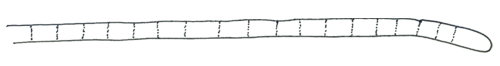
アルトロスピラ属 Arthrospira
プランクトトリックス属/プランクトトリコイデス属の項目を参照のこと。
プランクトトリックス属 Planktothrix/プランクトトリコイデス属 Planktothricoides
トリコームはまっすぐ、または緩く曲がるか規則的らせん形、幅は4μm以上で鞘はなく、分岐することはなく、単独で浮遊する。
細胞内全体にガス胞があり、チラコイドは放射状である。形態的な特徴だけではプランクトトリックス属 Planktothrixとプランクトトリコイデス属 Planktothricoidesとの区別は難しい。遺伝子配列(16S rRNA)の違いから両者は別属とされている。
一方、日本の淡水域に生育し、これまでArthrospira maxima(Spirulina maxima)またはArthrospira platensis(Spirulina platensis)と
同定されていた浮遊性種は、遺伝子解析の結果からプランクトトリックス属の1種Planktothrix spiroidesであることが明らかになっている。
Arthrospira maximaは主に熱帯や亜熱帯地域の塩水湖に生息し、形態および遺伝子解析の結果に基づき、現在はLimnospira属に移されている(Limnospira maxima)。またArthrospira platensisは淡水性の着生種である。
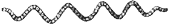
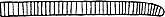
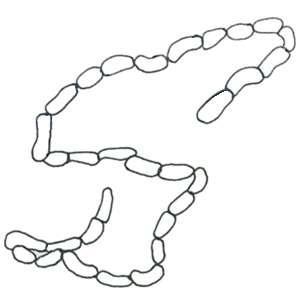
ユレモ属 Oscillatoria
トリコームはまっすぐか緩く曲がり、鞘はない、分枝はない、通常単独で生育することはなく、基質に着生し、目視できる程度の平らなマット状の群体を形成する。 止水域では、発達した群体全体が基質から離れ、水面に浮き上がることがある。細胞は円盤状で、幅約7μm以上、長さは幅の1/2から1/11、ガス胞はない。 日本で浮遊性のものはこれまでのところOscillatoria kawamuraeのみが知られている。
ミクロコレウス属 Microcoleus
藻体は鞘のないトリコームだけか、鞘内に一本のトリコームがあるか、または鞘内に多数のトリコームが含まれ、単独またはマット状に集合する。
トリコームはほぼまっすぐ、隔壁部でほとんどくびれず、先端部に向かってやや細くなる。細胞は円筒形、ガス胞はない。頂端細胞はやや細くなって丸く終わるか、円錐状に細くなり乳頭状に終わり、カリプトラがあることが多い。
本属の多くの種は湖岸や川岸などに着生し、運動性があるのに対し、不動の浮遊性種として報告されているのは現在のところM. pseudautumnalisだけである。
従来のミクロコレウス属のうち、トリコームの形態および遺伝子情報の違いからトリココレウス属Trichocoleusおよびコレオファシクルス属Coelophasciculusに移された種がある。
一方、フォルミディウム属Phormidiumのうち、一部の種は同じ理由からミクロコレウス属に移された。
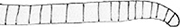
トリコデスミウム属 Trichodesmium
ユレモ型のトリコームが常に束を作って浮遊生活をする。T. iwanoffianum Nygaard と T. lacustre Kleb. が報告されている。

ネンジュモ目 Nostocales
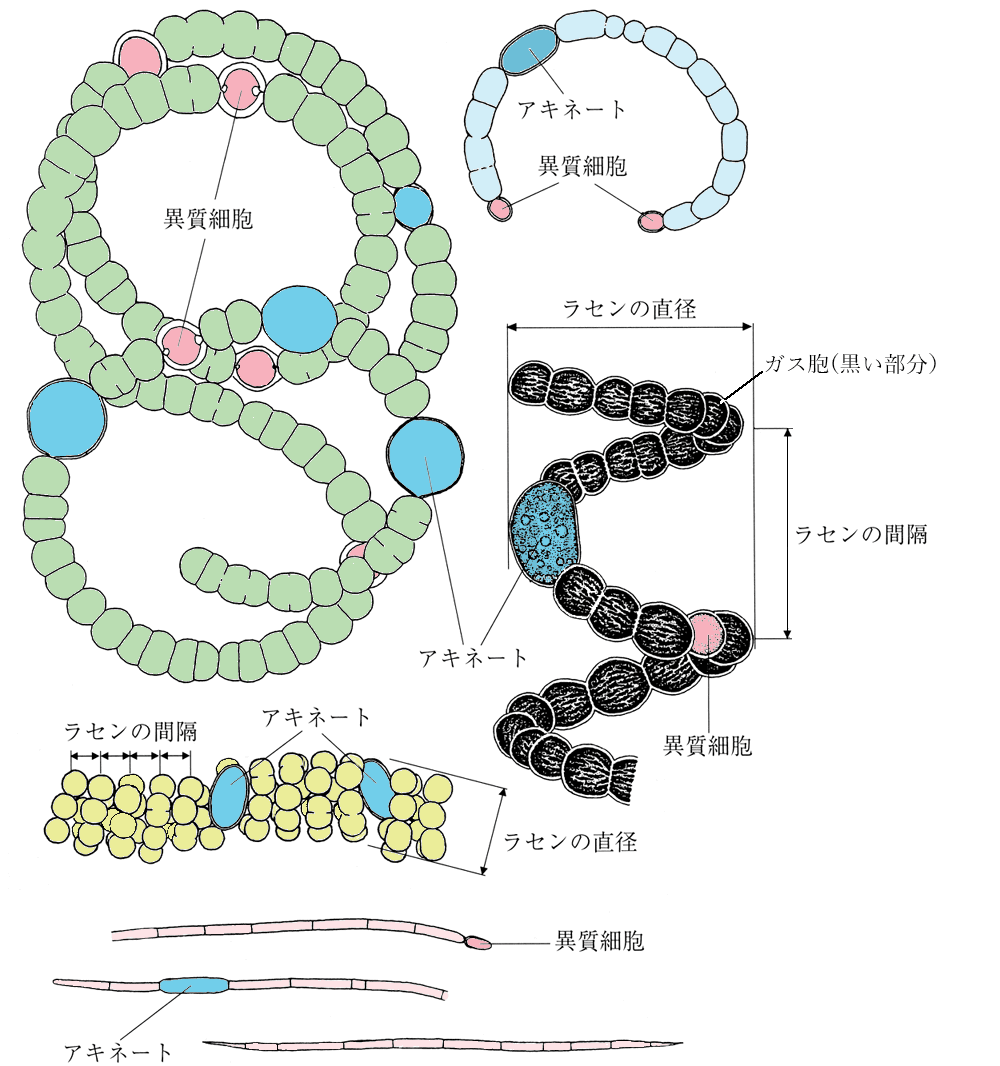
細胞は一列に並びトリコームを形成する。トリコームは栄養細胞、異質細胞、アキネートで構成される。しかし、トリコームが常に異質細胞やアキネートをもつわけではない。トリコームは分岐するかまたは分岐しない、鞘はあるかまたはない。
アナベノプシス属 Anabaenopsis
トリコームは単独で浮遊する。栄養細胞にはガス胞がある。トリコームの中央部の隣り合う2つの細胞がそれぞれ分裂し、隣接する側に形成された細胞が異質細胞に分化する。 したがって一時的にトリコームの中央部に異質細胞が2個並んで形成されるが、トリコームは2つの異質細胞が接する部位で分断するため、トリコームはその両端に異質細胞をもつことになる。 アキネートは楕円形または円筒形で栄養細胞より大きく、異質細胞から離れて1個または2個または数個連続してできる。 本邦ではA. arnoldii Aptekarおよび A. circinalis (G.S.West)Wołosz. et V.V.Mill.が報告されているが、大発生したという記録はない。
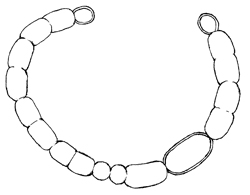
アファニゾメノン属 Aphanizomenon
トリコームは糸状で浮遊する、単独であることはまれで多数集合して束を形成する。トリコームはまっすぐかゆるく曲がり、全長を通じて同じ幅または先端部がやや細くなるが、先端は円いか平坦で針状にならない。 栄養細胞は円筒形、ガス胞がある、先端細胞はやや長い。異質細胞は円筒形または楕円形、栄養細胞とほぼ同じ幅である。アキネートは介生的、円筒形で栄養細胞より長く、異質細胞から離れて形成される。

クスピドスリクス属 Cuspidothrix
トリコームは糸状で単独で浮遊する。トリコームはまっすぐかゆるく曲がり、先端に向かって段階的に細くなり、尖って終わる。栄養細胞は円筒形、ガス胞があり、先端細胞は明らかに細く尖っている。 異質細胞は円筒形または楕円形、栄養細胞とほぼ同じ幅である。アキネートは介生的、円筒形で栄養細胞より長く、異質細胞から離れて形成される。 本邦では今のところC. issatschenkoi (Usačev) Rajaniemi et al.だけが報告されている。

以下にアファニゾメノン属(Aphanizomenon)とクスピドスリクス属(Cuspidothrix)の日本でみられる種について、形態の比較一覧表を示す。
▼表中の学名をクリックすると詳細をご覧になれます。
| 種名 | 群体の形態 | トリコームの形態 | トリコーム 先端細胞 |
アキネートの形態 | アキネートの長さ |
|---|---|---|---|---|---|
| A. flos-aquae | 大きな束状群体・紡錘形 | ほとんどまっすぐ | 長円筒形 | 長円筒形 | 約24-55μm |
| A. klebahnii | 小さな束状群体 | ほとんどまっすぐ | 円筒形 | 長円筒形 | 約18-31μm |
| A. paraflexuosum | 単独・群体をつくらない | ゆるく曲がる | 円筒形 | 長円筒形 | 約45-72μm |
| A. yezoense | 緩い束状群体 | ほとんどまっすぐ | 円筒形 | 長円筒形 | 約31-49μm |
| C. issatschenkoi | 単独・群体をつくらない | ほとんどまっすぐ | 先端細い針状 | 円筒形 | 約9-15μm |
キリンドロスペルモシス属 Cylindrospermopsis
ラフィディオプシス属 Raphidiopsisの項を参照のこと。
ドリコスペルマム属 Dolichospermum
トリコームはまっすぐか、規則的かやや不規則ならせん形、または不規則に曲がり絡み合っている。栄養細胞にはガス胞がある。異質細胞は球形または楕円形で、栄養細胞とほぼ同じ大きさで、トリコームの中間に形成される。 アキネートは球形、楕円形、円筒形で、幅も長さも栄養細胞や異質細胞より大きく、異質細胞の近くにできる。
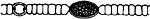
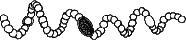
グロエオトリキア属 Gloeotrichia
トリコームは基部に異質細胞があり、先端部に向かって細くなり、毛状になって終わる。トリコームが異質細胞をもつ基部を中心にして放射状またはまばらに集合し、球形または半球形の群体をつくる。 アキネートは異質細胞に隣接する。本邦ではG. echinulata (J.E.Sm. et Soverby) P.G.Richit.が報告されている。

ラフィディオプシス属 Raphidiopsis
トリコームは単独で浮遊する。トリコームは比較的短く、直線状または円形またはらせん状に湾曲し、両端または一端が細く尖っており、鞘をもたない。異質細胞はトリコームの一端または両端にあるか、またはどちらにもない。アキネートはトリコームのほぼ中央部にできる。
従来、キリンドロスペルモプシス属 Cylindrospermopsisはトリコームの先端に異質細胞をつくるのに対し、ラフィディオプシス属は異質細胞をつくらないが、両属のトリコームやアキネートの形態などは差異がないといわれてきた。
しかし、両属の種が同一の水域に出現すること、トリコームが異質細胞をつくっていない場合はどちらの属なのか区別がつかないことなどから、成熟してアキネートと異質細胞の両方をもつものはキリンドロスペルモプシス属に、一方どちらもないかアキネートだけをもつものはラフィディオプシス属として扱われてきたのではないかとの疑問が以前からあった。
最近、形態だけでなく遺伝子情報(16S rRNA遺伝子、16S-23S rRNA ITS領域およびその二次構造)の解析が行われた結果、両属は単系統であり、別属として区別できないという研究報告があった(Aguirela et al. 2018)。この場合、植物命名規約の先取権の原則により、キリンドロスペルモプシス属の種はラフィディオプシス属に移されることになる。
本属は大発生してアオコをつくる場合がある。またRaphidiopsis raciborskii (シノニム:Cylindrospermopsis raciborskii)はシリンドロスパーモプシンやサキシトキシン、
Raphidiopsis curvispora
(シノニム:Cylindrospermopsis curvispora)はアナトキシンといった毒素をつくることが知られている。ただし、同一種でありながら毒素を産生しない培養株も報告されている。
スファエロスペルモプシス属 Sphaerospermopsis
トリコームはまっすぐか、または規則的かやや不規則ならせん形である。栄養細胞にはガス胞がある。異質細胞は栄養細胞とほぼ同じ大きさの球形で、トリコームの中間に形成される。 アキネートは球形で、栄養細胞や異質細胞より明らかに大きく、異質細胞の両側または片側に必ず隣接してできる。

以下にドリコスペルマム属(Dolichospermum)とスファエロスペルモプシス属(Sphaerospermopsis)の日本でみられる種について、形態の比較一覧表を示す。
▼表中の学名をクリックすると詳細をご覧になれます。
| 学名 | トリコームの形態 | 細胞の形態 | アキネートの形態 | 異質細胞とアキネートの位置関係 |
|---|---|---|---|---|
| D. affine | 直線状・束状群体 | 球形・樽型 | 楕円形・長楕円形 | 離れている |
| D. akankoense | 不規則らせん形 | レモン型 | 円筒形、多少曲がる | 近い |
| D. circinale | 規則的らせん形 | 球形・樽型 | 楕円形・円筒形 | 離れている |
| D. citrisporum | 直線状 | 球形・樽型 | レモン形 | 近い |
| D. crassum | 規則的らせん形 | 球形・樽型 | 広楕円形 | 離れている |
| D. flos-aquae | 不規則らせん形・絡み合う | 球形・樽型 | 円筒形、多少曲がる | 離れている |
| D. lemmermannii | 不規則らせん形・絡み合う | 球形・樽型 | 円筒形、多少曲がる | 隣接する |
| D. macrosporum | 直線状 | 球形・樽型 | 広楕円形・円筒形 | 離れている |
| D. mendotae | 不規則らせん形・絡み合う | 短円筒形 | 円筒形、多少曲がる | 離れている |
| D. minisporum | らせん形 | 球形・樽型 | 球形 | 離れている |
| D. mucosum | 不規則らせん形 | 球形・樽型 | 球形 | 離れている |
| D. planctonicum | 直線状 | 球形・樽型 | 広楕円形・円筒形 | 離れている |
| D. pseudocompactum | 密な規則的らせん形 | 球形・樽型 | 長楕円形、多少曲がる | 離れている |
| D. smithii | 直線状 | 球形・樽型 | 球形 | 離れている |
| D. spiroides | 規則的らせん形 | 球形・樽型 | 長楕円形、多少曲がる | 離れている |
| D. ucrainicum | 規則的らせん形 | 球形・樽型 | 球形 | 離れている |
| D. viguieri | 直線状 | 球形・樽型 | 広楕円形・円筒形 | 離れている |
| S. aphanizomenoides | 直線状 | 短円筒形・樽型 | 球形 | 隣接する |
| S. kisseleviana | 直線状 | 球形・樽型 | 球形 | 隣接する |
| S. oumiana | 規則的らせん形 | 球形・樽型 | 球形 | 隣接する |
| S. reniformis | 密な規則的らせん形 | 腎形・球形 | 球形 | 隣接する |
(新山・辻 2013より)
以下の種は日本での観察が限られており、上記の一覧表に含めていません。
- Anabaena cf. elliptica sensu M.Watan. 2007
- A. cf. heterospora sensu M.Watan. 2007
- A. cf. solitaria sensu M.Watan. 2006
- A. cf. tenericaulis sensu M.Watan. 2007
- A. levanderi Lemmerm. 1906
- A. tsugaruensis M.Watan. 2006
- Dolichospermum curvum (H. Hill) Wacklin et al. 2009
- Sphaerospermopsis eucompacta (R. Li et M.M.Watan.) X. Li et R. Li 2015
ウメザキア属 Umezakia
トリコームは単独で浮遊し、先端に向かってやや細くなる。栄養細胞にはガス胞があり、異質細胞とアキネートはトリコームの中間部に形成される。 培養中に分枝を形成したことから、当初スチゴネマ目で唯一の浮遊性種としてU. natans M.Watan. が報告された。 ところが、その後行われた遺伝子解析によって、本種がネンジュモ目の1グループに含まれること、しかしドリコスペルマム属、スファエロスペルモプシス属、クスピドスリクス属、ノデュラリア属などから明瞭に区別されることが明らかとなった。 形態的には、分枝がない状態では広義のアファニゾメノン属と、さらに異質細胞がなくアキネートをもつ状態ではラフィディオプシス属と区別することは難しい。今のところ、U. natans1種のみが知られており、日本以外からの報告はない。
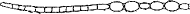

アエロトープ
ガス胞の集まり。
アキネート
トリコームをつくる栄養細胞が貯蔵物質を蓄積し、大型化し、厚い膜をつくり、生育に都合の悪い環境に耐えるために変化した細胞。耐久胞子あるいは単に胞子と呼ぶこともある。
異質細胞
トリコームをつくる栄養細胞が透明感のある黄緑色に変化して、厚い膜をつくったもの。窒素分子の好気的固定のために特別に分化した細胞と考えられている。ヘテロサイトともいう。シスト(胞嚢)ではないのでヘテロシストは間違い。
栄養細胞
光合成をして、2分裂によって増える普通の細胞のこと。
ガス胞
細胞内の微小な円筒形の小胞。浮遊性藍藻の浮力を調整している。ガス胞の膜はタンパク質で内部に気体を含有している。英語ではgas vesicleである。液胞とは異なるのでgas vacuoleは間違い。ガス胞の集まりをアエロトープという。
カリプトラ
糸状の藍藻のトリコームで、先端細胞の細胞壁が肥厚した構造。
チラコイド
薄膜状の細胞内構造。光合成反応に関わるクロロフィルaとフィコビリン色素を含んでいる。藍藻の分類では、細胞内のチラコイド配列の違いが重要と考えられている。
トリコーム
細胞が連続的に、糸状に並んだものを藍藻ではトリコームと呼ぶ。トリコームが周りに粘質物を分泌し、鞘に被われた状態のものを糸状体と呼びトリコームと区別する。